In search of the Holy Grail: Engineering the stem cell niche
26
SHARES
Posted: 19 April 2011 |
There is no other biomedical frontier that offers the stunning potential of human pluripotent stem cells and their progenitors in therapeutic applications to ease human suffering or in their ability to provide insights into development and diseases. Cell plasticity for reprogramming has revealed new opportunities in cell-based therapies and informed on lineage specification. What precisely defines each stem cell type or its transit amplifying progenitors that will lead to differentiated adult tissues is still being determined. Challenges remain in cell expansion, directed differentiation and environmental regulation of pluri- and multi-potent cells that avoid unwanted outcomes in transplantation therapies. Traditional culturing methods are giving way to a revolution in tissue engineering and biofabrication. The key to success is a multidisciplinary partnership of biologists, engineers, material scientists and clinicians. This strategy brings together cutting edge technologies and diverse expertise to bridge nano- to micro- to macroscale communication networks. Here, we discuss prominent technologies being applied to engineer the stem cell and tissue niche in vitro for the construction of 3D tissue architectures with integrated vascular networks.
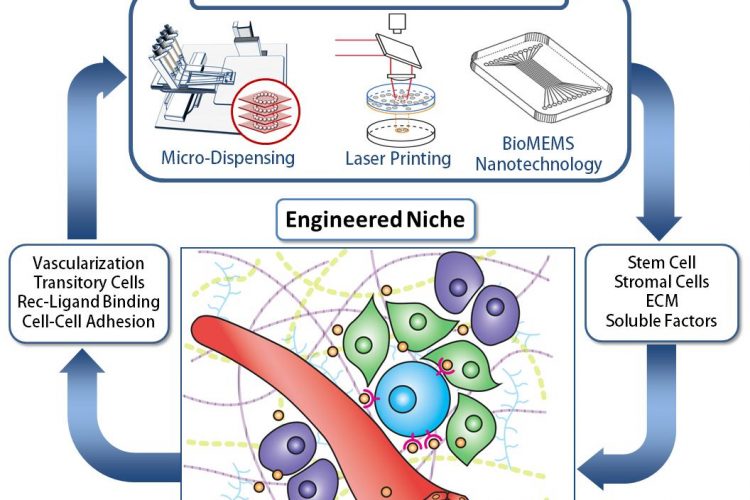
Figure 1. Engineering the Stem Cell Niche. The ability to assembly natural and manmade components into idealized biosynthetic 3D architectures requires a detailed understanding of the environment, along with the technology and expertise to manipulate them. This multiscale and multi-expertise approach is expected to lead to unprecedented breakthroughs in tissue engineering and biofabrication that benefit human health for an improved quality of life.
There is no other biomedical frontier that offers the stunning potential of human pluripotent stem cells and their progenitors in therapeutic applications to ease human suffering or in their ability to provide insights into development and diseases. Cell plasticity for reprogramming has revealed new opportunities in cell-based therapies and informed on lineage specification. What precisely defines each stem cell type or its transit amplifying progenitors that will lead to differentiated adult tissues is still being determined. Challenges remain in cell expansion, directed differentiation and environmental regulation of pluri- and multi-potent cells that avoid unwanted outcomes in transplantation therapies. Traditional culturing methods are giving way to a revolution in tissue engineering and biofabrication. The key to success is a multidisciplinary partnership of biologists, engineers, material scientists and clinicians. This strategy brings together cutting edge technologies and diverse expertise to bridge nano- to micro- to macroscale communication networks. Here, we discuss prominent technologies being applied to engineer the stem cell and tissue niche in vitro for the construction of 3D tissue architectures with integrated vascular networks.
This webinar showcases the Growth Direct System; an RMM (Rapid Microbial Method) that improves on traditional membrane filtration, delivering increased accuracy, a faster time to result, enhanced data integrity compliance, and more control over the manufacturing process.
Key learning points:
Understand the benefits of full workflow microbiology quality control testing automation in radiopharmaceutical production
Learn about ITM’s implementation journey and considerations when evaluating the technology
Find out how the advanced optics and microcolony detection capabilities of Growth Direct® technology impact time to result (TTR).
Don’t miss your chance to learn from experts in the industry –Register for FREE
Can’t attend live? No worries – register to receive the recording post-event.
The analysis of human cells in vitro provides invaluable insights into cell signalling and genomic stability and a source for protein production for human therapies. The estab – lishment of the first world resource for cells and tissues by the U.S. Navy in 1949 assisted routine culturing1,2. Almost 50 years later, a revolution in tissue culture and engineering took off in 1998 with the derivation of the first human embryonic stem cell (hESC) lines3. These pluripotent cells are derived from human blastocysts and are capable of differentiating into cells of all three germ layers. As such, they are expected to be unrivalled in their use in cell- or tissue-based therapies that improve the quality of human life4. Important information is also emerging from analysis of these cells in regard to human development and modelling of the earliest stages of human diseases5,6. Stem cells are, however, extremely complex. Over the past decade, our understanding of a stem cell at the level of transcriptional profiling, regulatory networks and epigenetic blueprints has evolved and is still being unravelled7-9. As the field of stem cell researchers expands, it now includes multidisciplinary teams that merge biology and clinical expertise with physics, engineering, materials science and mathematical modelling. Standardisation of protocols to identify, define and analyse pluripotent stem cells and their derivatives is underway and will optimise analyses and data compatibility that will advance safe therapeutic outcomes.
Promises and challenges
Untreatable, debilitating human diseases and injuries have found hope for treatment through the potential of stem cell therapies. While the need is urgent, ambitious clinical treatments without careful characterisation of transplanted stem cells has led to serious health consequences. In a treatment that brought international attention to stem cell therapies, neural stem cells injected into the brain of a young boy suffering from the incurable neurodegenerative disease Ataxia Telangiectasia resulted in a heterogeneous brain tumour originating from the donor tissue10. In response, international efforts have established resources that inform patients seeking stem cell therapies of the risks and need for validation of treatments. This includes the web resource established by the International Society of Stem Cell Research, ISSCR, entitled “A closer look at stem cell treatments”11.
The first stem cell therapies to use derivatives of pluripotent stem cells are now in clinical trials. Initiating trials are complicated in part by how to regulate these new agents, as devices or biologicals. Approved in 2009, and initiated in 2010, the California based company Geron, is using hESC derived donor cells to treat complete thoracic-level spinal cord injury in 10 patients in a phase 1 multicenter trial12. Oligodendrocyte progenitor cells derived from H1, one of the original hESC lines, will be evaluated for their ability to promote re-myelination and nerve growth in patients as modelled in mice13. How well mouse models will correlate with a human clinical outcome remains uncertain14. Differences in physiology and lifespan between animal models and humans limit the ability to fully predict contextual and long-term outcomes from animal models alone. An important question in transplantation is always whether the multidimensional tissue environment will provide additional unanticipated cues. This could lead to the transplanted cells entering a self-renewing tumorigenic cycle or straying from their intended lineage to generate alternative tissues or teratomas. Tumours arising from cancer stem cells occur, in part, through an inability of the environment to appropriately regulate self-renewal of these cells or their transient amplifying progenitors15-17. To drive human stem cell therapies, it is increasingly important to assess stem cell and progenitor cell potential in vitro.
The ability to mimic tissue and organ development and repair in xeno-free 2D or 3D architectures will allow modelling of multi-scale cellular events and tracking of transplanted cell fate. In addition, a better understanding of the stem cell niche, and ability to duplicate it in vitro, will remove other barriers to stem cell therapies that include expansion of pluripotent or multipotent cells in culture, avoidance of culture-derived artefacts and high efficiency directed differentiation of specific cell types.
Cellular reprogramming
Adult cells are often assumed to be irreversibly differentiated. In an exciting discovery by Takahashi and Yamanaka in 2006, embryonic like cells termed induced pluripotent stem (iPS) cells were derived from mouse embryonic and adult fibroblast cell populations using a transcription based approach18. Adult human fibroblasts can also be reprogrammed to a pluripotent state19,20, opening the door for patient specific stem cell therapies that could avoid immune rejection. IPS cells are also useful to model early developmental disease mechanisms that could lead to effective treatments6,7. Multiple cell types have now been successfully reprogrammed to iPS cells and alternative gene combination protocols tested21. Increasing the efficiency of iPS cell derivation, however, remains a challenge and must sidestep tumorigenicity and ageing defects that can be related to this technology22. Reprogramming strategies that alter the method of delivering the transcription signals, such as replacing viral transfection of genes with plasmid, mRNA or chemical based triggers appear to be successful in removing tumorigenic outcomes23. The most significant improvements to efficiency have come from environmental optimisation, such as using hypoxic growth to mimic physiologic states24-26. But are iPS cell lines equivalent to each other and to hESC lines? The pluripotent state shows differences at the level of transcriptome and epigenome between teratomas, hESC and iPSC lines27,28. IPS cell reprogramming can be incomplete, leaving epigenetic footprints that provide memory of previous lineage specification29. IPS cells also take time to derive, so hESCs or other banked stem cell lines, progenitor cells and stem cell derived tissues that have undergone standardised characterisation and validation protocols may be more readily available for stem cell based therapies.
Understanding how a multicellular organism emerges from a totipotent single celled embryo is enormously complex and underlies one of the challenges of working with cells in the pluripotent state.
A recent breakthrough in cell plasticity revealed an ability to partially reprogram adult somatic cells directly to intermediate multipotent progenitors. Direct conversion of fibroblasts to neurons in mouse models30 and the derivation of multipotent blood progenitors from human fibroblasts were demonstrated31. Strategies differed in each case. The direct reprogramming to neurons took advantage of chromatin remodelling and prominent transcription factors of the developing nervous system32-34, some of which are altered in human cancers35,36. The multipotent blood progenitors were derived by expression of the pluripotency transcription factor Oc4 followed by isolation of intermediate pan-hematopoietic CD45+ colonies that displayed a reduced pluripotency profile and down-regulated fibroblast specific genes. These cells transplanted into immunodeficient mice did not produce tumours, and the cells grown in culture were limited in cell divisions and not immortal37. Because the isolated therapeutic cell types may be limited in expansion in culture, it is important to optimise their efficacy for transplantation. The addition of supporting cells in 2D or 3D matrices with attention to impacts on genomic stability of these cells is needed38,39.
Multiscale modelling of the stem cell niche
Discoveries in the stem cell field are rapidly occurring due to the embracing of new disciplines in cell culturing, detailed profiling of cells, establishment of multidisciplinary teams and applications of cutting edge technologies. Greater attention to surface topology and biomaterials has brought nanotechnology rapidly into the arena of stem cell research. Nanoparticles are being used in real time biosensing to enable tracking of stem cells or progenitors in vivo, and as an intracellular delivery mechanism of cell regulators40. Choice of biomaterials is being combined with design of biomaterials, such as patterning of key protein substrates41 as well as precise nanopatterning of surfaces to promote material driven stem cell responses42. Nanoscale topography is expected to reveal better synthetic surfaces for cell growth, differentiation and multicellular patterning43. Indeed, effectively combining micro and nanostructured surfaces is revealing dramatic new insights in controlling cell orientation, adhesion and morphology that impact proliferation and differentiation44. How we apply nano and micro engineering technologies in in vitro tissue and organ modelling remains an exciting challenge45, one whose complexity will increasingly turn to multiscale modelling46.
The multifactorial challenge
Replicating signalling within the cellular niche is extremely challenging since it is composed of a multitude of interactions that includes cell-cell, cell-extracellular matrix (ECM), and cellbiochemical interactions (Figure 1)47. Understanding the influence of niche components and geometry is critical to regulating cell fate48. In order to understand the multi-factorial role of stem cell niche components in controlling stem cell fate, it must be deconstructed and synthetically reassembled in a controlled manner that can be iteratively modified by successively varying each motif of influence. As a simple yet powerful example, by varying the elastic moduli of the matrix substrate from 1 kPa to 10 kPa to 100 kPa one can regulate stem cell fate to neuronal, muscle or bone tissue, respectively49. What makes synthetic re-assembly of the cellular niche in an iterative manner most difficult is that the biomaterial components of tissue engineering are quite disparate in their manner of influence within the niche as well as the method of handling these biomaterials50-52.
Figure 1. Engineering the Stem Cell Niche. The ability to assembly natural and manmade components into idealized biosynthetic 3D architectures requires a detailed understanding of the environment, along with the technology and expertise to manipulate them. This multiscale and multi-expertise approach is expected to lead to unprecedented breakthroughs in tissue engineering and biofabrication that benefit human health for an improved quality of life.
Biofabrication
State-of-the-art tissue engineering currently involves homogeneously seeding cells into preformed scaffolds and exposing the construct to chemical or mechanical stimulation to achieve a desired fate. The materials used for these scaffolds are wide-ranging and include collagen sponges, silk, poly-L-lactide acid (PLLA) and porcine small intestine sub-mucosa to name a few. Their biocompatibility and amenability to heterogeneous cell type(s) and placement is limited in size and shape mostly due to the necessity of vasculature. The ability to build from the bottom up constructs that include heterogeneity in 3D and with all biomaterials would be a huge step towards reassembling the stem cell niche in vivo in a manner that best mimics the in vivo environment. Computer-aided design and computer-aided manufacturing (CAD/CAM) techniques like direct writing are becoming an increasingly important enabling tool towards this goal53.
Biofabrication can be defined as the use of CAD/CAM for the patterning and assembly of biomaterials54 (e.g., cells, scaffold and biochemicals) with a pre-defined 2D or even 3D organisation in order to produce idealised bioengineered constructs. Applications range from basic cell biology, pharmacology, to tissue engineering for regenerative medicine. The critical question throughout biofabrication is how relevant they are to mimicking the in vivo micro and nano-environments. The simplified schematic in Figure 1 (page 29) indicates the extreme complexity of the cellular niche. Many techniques are used in biofabrication, and this is not surprising since it is unlikely that any one technique can replicate the multitude of factors found within the in vivo cellular environment.
Controlled engineering the cellular niche began in 2D using material processing techniques common to electronics, e.g., photolithography. Using elastomeric stamps, 2D patterns of self-assembled monolayers of alkanethiols can be created as ~10 μm islands that support protein adsorption and cell attachment55. Patterns in poly(dimethylsiloxane) membranes have also been used to control cellular adherence properties as reflected in cell patterns on surfaces56. A more systematic study of nano-topography showed that corneal epithelial cells would elongate along groves in a substrate as small as 70 nanometres and concluded that topographic features in the basement membrane would have a similar effect57. This cellular response was shown to vary depending on the context in which the topographical stimuli were applied with different media, producing orthogonal responses58. Chen and coworkers recently used a micro-fabricated array of magnetic posts to demonstrate that cells respond to mechanical forces that are generated internally via their cytoskeleton59. Khademhosseini and coworkers summarised the use of microscale technologies for controlling the cellular microenvironment in vitro to direct cell fate60.
The different approaches to biofabrication can loosely be organised into four groups: twodimensional photolithographic patterning, dispensing techniques (e.g., ink jet or micropens)61,62, laser forward transfer63, and nano-dispensing64. Each technique has its own merits and shortcomings. We will expand hereafter on the capabilities of laser forward transfer as it is the most adept at engineering the cellular niche in terms of range of biomaterials to be deposited, spatial resolution, scalability, heterogeneity, amenability to 3D fabrication and assembly and accelerated tissue maturation. While there are a wide range of laser-based techniques for biofabrication, all the techniques that include living cell deposition can be termed laser forward transfer techniques. In what follows, we briefly summarise the demonstration of the biofabrication of 3D constructs of the biomaterials relevant to the stem cell niche and tissue engineering.
The proximity of cells in culture greatly affects cellular behaviour either through direct cell-cell contacts or indirectly through paracrine signalling. Sufficiently large cellular distances require communication through endocrine signalling. Thus, in addition to the geometric constraints on growth and/or migration imposed by neighbouring cells, cell spacing in a synthetic niche will affect the manner in which the cells communicate in culture. This is of particular interest when culturing pluripotent cells, since environmental cues and signalling pathways influence their differentiation and ultimate fate. In stem cell cultures, cell spatial arrangements contribute to cell fate choices. When cultured on collagen, human mesenchymal stem cells arranged in a confluent monolayer differentiated into an osteoblast lineage, whereas the same cells cultured in small clusters developed into chondroblasts65. Relative cell density in culture appears to be less critical in a study of rat mesenchymal stem cell multilineage potential66. Cellular spacing within the more complex in vivo niche environments in maintenance and differentiation has yet to be fully quantified or characterised.
In addition to cellular proximity, neigh – bouring cells have a profound influence on cell behaviour via intercellular communication. Recent co-culturing studies with human mesenchymal stem cells reveal the importance of neighbouring cell types on redirecting protein expression profiles67-69. Pedrotty et al. showed that cardiomyoctyes displayed different protein expression patterns when co-cultured with human mesenchymal stem cells than when cultured homotypically. Furthermore, the type of expression was location dependent, i.e., the expression was altered from purely junctions (at cell-cell contact) in homotypic cardiomyocyte pairs, to a mixture of junction and diffuse (inside the cytoplasm) for the heterotypic co-culture. These recent findings highlight the importance of choosing appropriate neighbouring cell types in directing stem cell differentiation. It also underscores the importance of investigating not only changes to levels of expression, but also the cellular mechanisms affected. Finally, the most advanced generation of 3D tissue architectures will require integration of neural and vascular networks. Here we discuss challenges with engineering in the vascular niche.
The vascular niche
The importance of the vascular niche in work with stem cells is increasingly evident and was found to be critical in controlling stem cell differentiation within multiple tissues and organs. For example, endothelial cells (ECs) are essential for the self-renewal and repopulation of hematopoietic stem cells70,71. In the nervous system, adult neural stem cells reside in a vascular niche72-74. In addition, the vascular niche has been associated with many other tissue-specific stem cells such as cardiac stem cells75,76, adipose tissue derived stem cells77 and cancer stem cells78,79. Engineering stem cells along these lineages in vitro, for research or therapeutic purposes, has had limited success due to the difficulty to accurately recreate the in vivo niche, even in the presence of necessary cytokines and serum. The vascular niche resides in an anatomically defined, specifically constituted place. Many signals from the niche are involved in controlling stem cell fate, including biochemical stimuli, cell-cell interactions and cell-matrix inter actions80. In addition, vascular endothelial cells are constantly subjected to shear stress and stretch caused by the flowing blood and pulsatile pressure. Mimicking this complex 3D micro – environment in vitro has been a major challenge in vascular research. It is apparent that traditional cell culture techniques and co-culture models are not sufficient to mimic the complex architecture of the vascular niches. Therefore, developing new technologies is critical to overcome this challenge. In the past decade, emerging technologies in the field of microfabrication, tissue engineering and biomaterials have been developed, which provide new approaches to address vascularisa tion. Here we will review some of the most recent developments in this field and its application in stem cell research.
Engineering the vascular niche
Methods to engineer vascular networks are generally conducted following two main approaches. The first approach relies primarily on vascular cells’ innate ability to regenerate. The cells themselves provide the necessary signalling cues that allow the vascular cells to organise into a functional vascular network. In the past 20 years, there has been tremendous progress in the development of novel biomaterial scaffolds with tailored biochemical and biomechanical properties81. Technologies have also been developed to create 3D architectures with micro- to nano- level features to mimic certain advantageous characteristics of the natural extracellular matrix for vascular generation82. Other approaches involve embedding various angiogenesis growth factors within the engineered matrix for controlled release to promote endothelial cell invasion into the construct after implantation83-85.
The second approach to engineer vascular networks involves designing and fabricating vascular channels mimicking blood vessel networks using novel engineering technologies. Nanoscale and microfabrication technologies, the same set of techniques used within the semiconductor industry, have been introduced into the field of tissue engineering. With these tools, one can easily create any pattern of proteins, extracellular matrix and cells on 2D surfaces at very high resolution (nm~μm). This allows precise control of cell shape and cell-cell interactions86,87, which are invaluable for investigating specific requirements of the stem cell niche88-91. Microfabrication technology has also been employed to create arrays microchannels in biodegradable polymers or hydrogels. Endothelial cells are then seeded into the channels for the formation of vascular network92-98. Recently, a manufacturing technique commonly used in industry, known as 3D free-form fabrication (3DFFF), has been successfully applied to tissue engineering by fabricating complex hierarchical biomaterial scaffolds99-109. This technology allows on-demand control of cells, growth factors and matrix in predefined configuration to mimic tissue’s natural structures, and has been implemented to create vascular channels110-114. Although still at an early stage, enabling technologies in the field of tissue engineering have begun to show significance in stem cell research.
Future perspectives The future of tissue engineering for cell-based therapies to increase the quality of human life is an exciting and obtainable frontier. However, major obstacles and challenges still need to be addressed. Many of the current technologies, although powerful for their particular applications (e.g., generating a particular niche), are not generally transferable. Simultaneous incorporation of multiple niche components (cell types, ECM, biomechanical stimuli, growth factors; Figure 1 (page 29) is still cumbersome. For vascular network formation, going from a 2D network array to complex 3D vasculature still faces tough challenges. Many of the technologies discussed here involve sophisticated instrumentation and software as well as a complex material fabrication process, making them not readily accessible to the majority of the stem cell researchers. Only through establishing close collaborations between skilled researchers in different areas of technological, biological and materials expertise can tissue engineering with stem cells move rapidly forward.
Acknowledgements
This work is supported by NYSTEM C026186 (JLP); R21HL102773 (GD); and NIH R56- DK088217 (DBC).
The authors thank graduate student Vivian Lee for her contribution to the final design of Figure 1.
About the Authors
Janet L. Paluh is an Associate Professor in Nanobioscience in the College of Nanoscale Science and Engineering at the University at Albany, SUNY and Chair of the Capital region Stem Cell Oversight Committee. She received her PhD in Cancer Biology from Stanford University and completed her post-doctoral training at the University of California, Berkeley. She served as a research Assistant Professor at Rensselaer Polytechnic Institute before joining CNSE. Her research applies single and multicellular investigation with nanoengineering and nanomedicine. Research interests include genomic stability, pluripotent stem cells and tissue modelling in 3D and biosensing as well as analysis of molecular machines in multi-scale communication networks from nano-, micro- to macroscale.
Guohao Dai is an Assistant Professor in the Department of Biomedical Engineering at Rensselaer Polytechnic Institute. He received his PhD in Biomedical Engineering from Massachusetts Institute of Technology, and completed postdoctoral training at Harvard Medical School. Guohao’s research interests are in cardiovascular biomechanics and vascular bioengineering.
Douglas B. Chrisey is a Professor of Materials Science and Biomedical Engineering at Rensselaer Polytechnic Institute. He received his PhD in Physics from the University of Virginia in 1987 and then joined the US Naval Research Laboratory where he spent the next 17 years. His current research interests include the novel laser fabrication of thin films and coatings for advanced electronics, sensors, biomaterials and energy storage. These materials were used in device configurations for testing and typically have an improved figure-of-merit. He is considered one of the pioneers in the field of Pulsed Laser Deposition and was the lead inventor of MAPLE processing technique. His research has resulted in more than 300 citable publications and over 7,000 citations and he has edited or co-edited 15 books and has 18 patents. He also serves as a Scientific Advisor to Nanotherapeutics, the Chief Technology Officer for Nano Solutions and is President of Omni-Metrics.
References
1. Stacey GN, Masters JR. Cryopreservation and banking of mammalian cell lines. Nat Protoc, 3: 1981-1989 (2008)
2. Strong DM. The US Navy Tissue Bank: 50 years on the cutting edge. Cell and Tissue Banking, 1: 9-16 (2000)
3. Thomson JA, Itskovitz-Eldor J, Shapiro SS, Waknitz MA, Swiergiel JJ, Marshall VS, Jones JM. Embryonic stem cell lines derived from human blastocysts. Science, 282: 1145- 1147 (1998)
5. Keller G. Embryonic stem cell differentiation: emergence of a new era in biology and medicine. Genes Dev, 19: 1129-1155 (2005)
6. Lee G, Papapetrou EP, Kim H, Chambers SM, Tomishima MJ, Fasano CA, Ganat YM, Menon J, Shimizu F, Viale A, Tabar V, Sadelain M, Studor L. Modeling pathogenesis and treatment of familial dysautonomia using patient specific iPSCs. Nature, 461: 402-406 (2009)
7. Bibikova M, Laurent LC, Ren B, Loring JF, Fan J-B. Unraveling epigenetic regulation in embryonic stem cells. Cell Stem Cell, 2: 123-134 (2008)
8. Li H, Liu Y, Shin S, Sun Y, Loring JF, Mattson MP, Rao MS, Zhan M. Transcriptome coexpression map of human embryonic stem cells. BMC genomics, 7: 103-117 (2006)
9. Müller F-J, Laurent LC, Kostka D, Ulitsky I, Williams R, Lu C, Park I-H, Rao MS, Shamir R, Schwartz PH, Schmidt NO, Loring JF. Regulatory networks define phenotypic classes of human stem cell lines. Nature, 455: 401-406 (2008)
10. Amariglio N, Hirshberg A, Scheithauer BW, Cohen Y, Loewenthal R, Trakhtenbrot L, Paz N, Koren-Michowitz M, Waldman D, Leider-Trejo L, Toren A, Constantini S, Rechavi G. Donor derived brain tumor following neural stem cell transplantation in an ataxia telangiectasia patient. PLoS Med, 6: e1000029 (2009)
12. Alper J. Geron gets green light for human trial of ES cellderived product. Nat. Biotech, 27: 213-214 (2009)
13. Keirstead HS, Nistor G, Bernal G, Totoin M, Cloutier F, Sharp K, Steward O. Human embryonic stem cell-derived oligodendrocyte progenitor cell transplants remyelinate and restore locomotion after spinal cord injury. J Neurosci, 25: 4694-4705 (2005)
14. Greber B, Wu G, Bernemann C, Joo JY, Han DW, Ko K, Tapia N, Sabour D, Sterneckert J, Tesar P, Schöler HR. Conserved and divergent roles of FGF signaling in mouse epiblast stem cells and human embryonic stem cells. Cell Stem Cell, 6: 215-226 (2010)
15. Tan BT, Park CY, Ailles LE, Weissman IL. The cancer stem cell hypothesis: a work in progress. Lab. Invest, 86: 1203- 1207 (2006)
16. Sell S. On the stem cell origin of cancer. Am. J. Pathol, 176: 2584-2594 (2010)
17. Reya T, Morrison SJ, Clarke MF, Weissman, IL. Stem cells, cancer, and cancer stem cells. Nature, 414: 105-111 (2001)
18. Takahashi K, Yamanaka S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell, 126: 663-676 (2006)
19. Takahashi K, Tanabe K, Ohnuki M, Narita M, Ichisaka T, Tomoda K, Yamanaka S. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell, 131: 861-872 (2007)
20. Yu J, Vodyanik MA, Smuga-Otto K, Antosiewicz-Bourget J, Frane JL, Tian S, Nie J, Jonsdottir GA, Ruotti V, Stewart R, Slukvin II, Thomson JA. Induced pluripotent stem cell lines derived from human somatic cells. Science 318: 1917-1920 (2007)
21. Rodolfa KT, Eggan K. A transcriptional logic for nuclear reprogramming. Cell, 126: 652-655 (2006)
22. Okita K, Yamanaka S. Induction of pluripotency by defined factors. Exp. Cell Res, 316: 2565-2570 (2010)
23. Jaenisch R, Young R. Stem cells, the molecular circuitry of pluripotency and nuclear reprogramming. Cell, 132: 567-582 (2008)
24. Csete M. Oxygen in the cultivation of stem cells. Ann NY Acad Sci, 1049:1-8 (2005)
25. Yoshida Y, Takahashi K, Okita K, Ichisaka T, Yamanaka S. Hypoxia enhances the generation of induced pluripotent stem cells. Cell Stem Cell, 5: 237-241 (2009)
26. Mohyeldin, A., Garzón-Muvdi, T., Quiñones-Hinojosa, A. Oxygen in stem cell biology: a critical component of the stem cell niche. Cell Stem Cell 2010, 7, 150-161(2010)
27. Bibikova M, Chudin E, Wu B, Zhou L, Garcia EW, Liu Y, Shin S, Plaia TW. et al. Human embryonic stem cells have a unique epigenetic signature. Genome Res, 16: 1075-1083 (2006)
28. Marchetto MCN, Yeo GW, Kainohana O, Marsala M, Gage FH, Muotri AR. Transcriptional signature and memory retention of human-induced pluripotent stem cells. PLoS One, 4: e7076 (2009)
29. Kim K, Doi A, Wen B, Ng K, Zhao R, Cahan P et al. Epigenetic memory in induced pluripotent stem cells. Nature, 467: 285-290 (2010)
30. Vierbuchen T, Ostermeier A, Pang ZP, Kokubu Y, Südhof TC, Wernig M. Direct conversion of fibroblasts to functional neurons by defined factors. Nature, 463: 1035- 1042 (2010)
31. Szabo E, Rampalli S, Risueño RM, Schnerch A, Mitchell R, Fiebig-Comyn A, Levadoux-Martin M, Bhatia M. Direct conversion of human fibroblasts to multilineage blood progenitors. Nature, 468: 521-526 (2010)
32. Romm E, Nielsen JA, Kim JG, Hudson LD. Myt1 family recruits histone deacetylase to regulate neural transcription. J Neurochem, 93: 1444-1453 (2005)
33. Ryan AK, Rosenfeld MG. Pou domain family values: flexibility, partnerships, and developmental codes. Genes Dev, 11: 1207-1225 (1997)
34. Kageyama R, Ishibashi M, Takebayashi K, Tomita K. bHLH transcription factors and mammalian neuronal differentiation. Int J Biochem Cell Biol, 29: 1389-1399 (1997)
35. Ball DW. Achaete-scute homolog-1 and Notch in lung neuroendocrine development and cancer. Cancer Letters, 204: 159-169 (2004)
36. Cook AL, Sturm RA. POU domain transcription factors: BRN2 as a regulator of melanocytic growth and tumourigenesis. Pigment Cell Melanoma Res, 21: 611-626 (2008)
37. Lebofsky R, Walter JC. New Myc-anisms for DNA replication and Tumorigenesis? Cancer Cell, 12: 102- 103 (2007)
38. Maitra A, Arking DE, Shivapurkar N, Ikeda M, Stastny V, Kassauei K, Sui G, Cutler DJ, Liu Y, Brimble SN, Noaksson K, Hyllner J, Schulz TC, Zeng X, Freed WJ, Crook J, Abraham S, Colman A, Sartipy P, Matsui S-I, Carpenter M, Gazdar AF, Rao M, Chakravarti A. Genomic alterations in cultured human embryonic stem cells. Nat Genet, 37: 1099-1103 (2005)
39. Joseph NM, Morrison SJ. Toward an understanding of the physiological function of mammalian stem cells. Develop Cell, 9: 173-183 (2006)
40. Ferreira L. Nanoparticles as tools to study and control stem cells. J Cell Bioch, 108: 746-752 (2009)
41. Lutolf MP, Gilbert PM, Blau HM. Designing materials to direct stem cell fate. Nature, 462: 433-441 (2009)
42. Curran JM, Chen R, Stokes R, Irvine E, Graham D, Gubbins E, Delaney D, Amro N, Sanedrin R, Jamil H, Hunt JA. Nanoscale definition of substrate materials to direct human adult stem cells towards tissue specific populations. J Mater Sci Mater Med, 21: 1021-1029 (2010)
44. Martinez E, Engel E, Planell JA, Samitier J. Affects of artificial micro- and nano-structured surfaces on cell behaviour. Ann Anat, 191: 126-135 (2009)
45. Willerth SM, Sakiyama-Elbert SE. Combining stem cells and biomaterial scaffolds for constructing tissues and cell delivery. In StemBook [internet], Cambridge (MA), Harvard stem cell institute. pp. 1-18 (2008)
46. Engler AJ, Humbert PO, Wehrle-Haller B, Weaver VM. Multiscale modeling of form and function. Science, 324: 208-212 (2009)
47. Jordan JD, Landau EM, and Lyengar R. Signaling networks: The origins of cellular multitasking. Cell 103: 193-200 (2000)
48. Morrison SJ and Spradling AC. Stem cells and niches: Mechanisms that promote stem cell maintenance throughout life. Cell 132: 598-611 (2008)
49. Even-Ram S, Artym V, and Yamada KM. Matrix control of stem cell fate. Cell 126:645-647 (2006)
50. Teixeira AI, Hermanson O, and Werner C. Designing and engineering stem cell niches. MRS Bulletin 35: 591-596 (2010)
51. Vazin T and Schaffer DV. Engineering strategies to emulate the stem cell niche. Trends in Biotechnology 28: 117-124 (2010)
52. Peerani R and Zandstra PW. Enabling stem cell therapies through synthetic stem cell-niche engineering. Journal of Clinical Investigation 120:60-70 (2010)
53. Chrisey DB. Material processing – The power of direct writing. Science 289:879-881 (2000)
54. Pique A and Chrisey DB. Direct-Write Technologies for Rapid Prototyping: Applications to Electronics, Sensors, and Passivation Coatings. (2002) Academic Press, New York (2004)
55. Singhvi R, Kumar A, Lopez G, Stephanopoulos GN, Wang DIC, Whitesides GM, and Ingber DE. Engineering Cell Shape and Function. Science 264:696-698 (1994)
56. Ostuni E, Kane R, Chen CS, Ingber DE, and Whitesides GM. Patterning mammalian cells using elastomeric membranes. Langmuir 16:7811-7819 (2000)
57. Teixeira AI, Abrams GA, Bertics PJ, Murphy CJ, and Nealey PF. Epithelial contact guidance on well-defined micro and nanostructured substrates. J. of Cell Science 116:1881- 1892 (2003)
58. Teixeira AI, McKie GA, Foley JD, Berticsc PJ, and Murphy CJ. The effect of environmental factors on the response of human corneal epithelial cells to nanoscale substrate topography. Biomaterials 27:3945-3954 (2006)
59. Sniadecki NJ, Anguelouch A, Yang MT, Lamb CM, Liu Z, Kirschner SB, Liu Y, Reich DH, and Chen CS. Magnetic microposts as an approach to apply forces to living cells. Proceedings of the National Academy of Science 104:14553-14558 (2007)
60. Khademhosseini A, Langer R, Borenstein J, and Vacanti JP. Microscale technologies for tissue engineering and biology. Proceedings of the National Academy of Sciences 103:2480-2487 (2006)
61. Im M, Cho IJ, Yun KS, and Yoon E. Electromagnetic actuation and microchannel engineering of a polymer micropen array integrated with microchannels and sample reservoirs for biological assay patterning. Applied Physics Letters 91:124101 (2007)
62. Pardo L, Wilson WC, and Boland TJ. Characterization of patterned self-assembled monolayers and protein arrays generated by the ink-jet method. Langmuir 19:1462- 1466 (2003
63. Schiele NR, Koppes RA, Corr DT, Ellison KS, Thomposon DM, Ligon LA, Lippert TKM, and Chrisey DB. Laser direct writing of combinatorial libraries of idealized cellular constructs: Biomedical applications. Applied Surface Science 255:5444-5447 (2009)
64. Piner RD, Zhu J, Xu F, Hong SH, and Mirkin CA. Dip-pen nanolithography. Science 283:661-663 (1999)
65. Martin BJ, Senechai G, Pittenger MF, and Byrne BJ. Circulation 100:54 (1999)
66. Neuhuber B, Swanger SA, Howard L, Mackay A, and Fischer I. Effects of plating density and culture time on bone marrow stromal cell characteristics. Experimental Hematology 36:1176-1185 (2008)
67. Vadala G, Sobajima S, Lee JY, Huard J, Denaro V, Kang JD, and Gilbertson LG. Spine Journal 8:804-809 (2008)
68. Xu Y, Shi Y, and Ding S. A chemical approach to stem-cell biology and regenerative medicine. Nature 453: 338-344 (2008)
69. Pedrotty DM, Klinger RY, Badie N, Hinds S, Kardashian A, and Bursac N. Structural coupling of cardiomyocytes and noncardiomyocytes: quantitative comparisons using a novel micropatterned cell pair assay. American Journal of Physiology – Heart and Circulartory Physiology 295:H390-H400 (2008)
70. Butler JM, Nolan DJ, Vertes EL, Varnum-Finney B, Kobayashi H, Hooper AT, Seandel M, Shido K, White IA, Kobayashi M, et al. Endothelial cells are essential for the self-renewal and repopulation of Notchdependent hematopoietic stem cells. Cell stem cell 6: 251-264 (2010)
71. Wilson A, and Trumpp A. Bone-marrow haematopoieticstem- cell niches. Nature Reviews Immunology 6: 93-106 (2006)
72. Shen Q, Wang Y, Kokovay E, Lin G, Chuang S-M, Goderie SK, Roysam B, and Temple S. Adult SVZ stem cells lie in a vascular niche: a quantitative analysis of niche cell-cell interactions. Cell Stem Cell 3: 289-300 (2008)
73. Shen Q, Goderie SK, Jin L, Karanth N, Sun Y, Abramova N, Vincent P, Pumiglia K, and Temple S. Endothelial cells stimulate self-renewal and expand neurogenesis of neural stem cells. Science 304: 1338-1340 (2004)
74. Tavazoie M, Van der Veken L, Silva-Vargas V, Louissaint M, Colonna L, Zaidi B, Garcia-Verdugo JM, and Doetsch F. A specialized vascular niche for adult neural stem cells. Cell Stem Cell 3: 279-288 (2008)
75. Anversa P, Kajstura J, Leri A, and Bolli R. Life and death of cardiac stem cells: a paradigm shift in cardiac biology. Circulation 113: 1451-1463 (2006)
76. Leri A, Kajstura J, and Anversa P. Cardiac stem cells and mechanisms of myocardial regeneration. Physiological reviews 85: 1373-1416 (2005)
77. Tang W, Zeve D, Suh JM, Bosnakovski D, Kyba M, Hammer RE, Tallquist MD, and Graff JM. White fat progenitor cells reside in the adipose vasculature. Science 322: 583-586 (2008)
78. Calabrese C, Poppleton H, Kocak M, Hogg TL, Fuller C, Hamner B, Oh EY, Gaber MW, Finklestein D, Allen M, et al. A perivascular niche for brain tumor stem cells. Cancer Cell 11: 69-82 (2007)
79. Gilbertson RJ, and Rich JN. Making a tumour’s bed: glioblastoma stem cells and the vascular niche. Nat Rev Cancer 7: 733-736 (2007)
80. Morrison SJ, and Spradling AC. Stem cells and niches: mechanisms that promote stem cell maintenance throughout life. Cell 132: 598-611 (2008)
81. Langer R, and Tirrell DA. Designing materials for biology and medicine. Nature 428: 487-492 (2004)
82. Ma PX. Biomimetic materials for tissue engineering. Adv Drug Deliv Rev 60:184-198 (2008)
83. Richardson TP, Peters MC, Ennett AB, and Mooney DJ. Polymeric system for dual growth factor delivery. Nat Biotechnol 19: 1029-1034 (2001)
84. Lee KY, Peters MC, Anderson KW, and Mooney DJ. Controlled growth factor release from synthetic extracellular matrices. Nature 408: 998-1000 (2000)
85. Lutolf MP, and Hubbell JA. Synthetic biomaterials as instructive extracellular microenvironments for morphogenesis in tissue engineering. Nat Biotechnol 23: 47-55 (2005)
86. Chen CS, Mrksich M, Huang S, Whitesides GM, and Ingber DE. Geometric control of cell life and death. Science 276: 1425-1428 (1997)
87. Bhatia SN, Yarmush ML, and Toner M. Controlling cell interactions by micropatterning in co-cultures: hepatocytes and 3T3 fibroblasts. J Biomed Mater Res 34: 189-199 (1997)
88. Khademhosseini A, Langer R, Borenstein J, and Vacanti JP. Microscale technologies for tissue engineering and biology. Proc Natl Acad Sci U S A 103: 2480-2487 (2006)
89. Liu WF, and Chen CS. Cellular and multicellular form and function. Adv Drug Deliv Rev 59: 1319-1328 (2007)
90. Khetani SR, and Bhatia SN. Engineering tissues for in vitro applications. Curr Opin Biotechnol 17: 524-531 (2006)
91. Kane RS, Takayama S, Ostuni E, Ingber DE, and Whitesides GM. Patterning proteins and cells using soft lithography. Biomaterials 20: 2363-2376 (1999)
92. Borenstein JT, Tupper MM, Mack PJ, Weinberg EJ, Khalil AS, Hsiao J, and García-Cardeña G. Functional endothelialized microvascular networks with circular cross-sections in a tissue culture substrate. Biomed Microdevices 12: 71-79 (2010)
93. Shin M, Matsuda K, Ishii O, Terai H, Kaazempur-Mofrad M, Borenstein J, Detmar M, and Vacanti JP. Endothelialized networks with a vascular geometry in microfabricated poly(dimethyl siloxane). Biomed Microdevices 6: 269-278 (2004)
94. Chung S, Sudo R, Vickerman V, Zervantonakis IK, and Kamm RD. Microfluidic platforms for studies of angiogenesis, cell migration, and cell-cell interactions. Sixth International Bio-Fluid Mechanics Symposium and Workshop March 28-30, Pasadena, California. Ann Biomed Eng 38: 1164-1177 (2010)
95. King K, Wang C, Kaazempur Mofrad M, Vacanti J, and Borenstein J. Biodegradable microfluidics. Advanced Materials 16: 2007-2012 (2004)
96. Bettinger C, Weinberg E, Kulig K, Vacanti J, Wang Y, Borenstein J, and Langer R. Three Dimensional Microfluidic Tissue Engineering Scaffolds Using a Flexible Biodegradable Polymer. Advanced Materials 18: 165-169 (2006)
97. Golden AP and Tien J. Fabrication of microfluidic hydrogels using molded gelatin as a sacrificial element. Lab Chip 7: 720-725 (2007)
98. Choi NW, Cabodi M, Held B, Gleghorn JP, Bonassar LJ, and Stroock AD. Microfluidic scaffolds for tissue engineering. Nat Mater 6: 908-915 (2007)
99. Dhariwala B, Hunt E, and Boland T. Rapid prototyping of tissue-engineering constructs, using photopolymerizable hydrogels and stereolithography. Tissue Eng 10: 1316-1322 (2004)
100. Cooke MN, Fisher JP, Dean D, Rimnac C, and Mikos AG. Use of stereolithography to manufacture critical-sized 3D biodegradable scaffolds for bone ingrowth. J Biomed Mater Res B Appl Biomater 64: 65-69 (2003)
101. Yang S, Leong KF, Du Z, and Chua CK. The design of scaffolds for use in tissue engineering. Part II. Rapid prototyping techniques. Tissue Eng 8: 1-11(2002)
102. Hollister SJ. Porous scaffold design for tissue engineering. Nat Mater 4: 518-524 (2005)
103. Peltola SM, Melchels FP, Grijpma DW, and Kellomaki M. A review of rapid prototyping techniques for tissue engineering purposes. Ann Med 40: 268-280 (2008)
104. Sun W, Darling A, Starly B, and Nam J. Computer-aided tissue engineering: overview, scope and challenges. Biotechnol Appl Biochem 39: 29-47 (2004)
105. Yeong WY, Chua CK, Leong KF, and Chandrasekaran M. Rapid prototyping in tissue engineering: challenges and potential. Trends Biotechnol 22: 643-652 (2004)
106. Mironov V, Visconti RP, Kasyanov V, Forgacs G, Drake CJ, and Markwald RR. Organ printing: tissue spheroids as building blocks. Biomaterials 30: 2164-2174 (2009)
107. Roth EA, Xu T, Das M, Gregory C, Hickman JJ, and Boland T. Inkjet printing for high-throughput cell patterning. Biomaterials 25: 3707-3715 (2004)
108. Xu T, Jin J, Gregory C, Hickman JJ, and Boland T. Inkjet printing of viable mammalian cells. Biomaterials 26: 93-99 (2005)
109. Lee W, Debasitis JC, Lee VK, Lee JH, Fischer K, Edminster K, Park JK, and Yoo SS. Multi-layered culture of human skin fibroblasts and keratinocytes through three-dimensional freeform fabrication. Biomaterials 30: 1587-1595 (2009)
110. Norotte C, Marga FS, Niklason LE, and Forgacs G. Scaffoldfree vascular tissue engineering using bioprinting. Biomaterials 30: 5910-5917 (2009)
111. Cui X, and Boland T. Human microvasculature fabrication using thermal inkjet printing technology. Biomaterials 30: 6221-622 (2009)
112. Lee W, Lee V, Polio S, Keegan P, Lee J-H, Fischer K, Park J-K, and Yoo S-S. On-demand three-dimensional freeform fabrication of multi-layered hydrogel scaffold with fluidic channels. Biotechnol Bioeng 105: 1178-1186 (2010)
113. Nahmias Y, Schwartz RE, Verfaillie CM, and Odde DJ. Laser-guided direct writing for three-dimensional tissue engineering. Biotechnol Bioeng 92: 129-136 (2005)
114. Schiele NR, Corr DT, Huang Y, Nurazhani AR, Xie Y, and Chrisey DB. Laser-based direct-write techniques for cell printing. Biofabrication 2: 032001 (2010)
This website uses cookies to enable, optimise and analyse site operations, as well as to provide personalised content and allow you to connect to social media. By clicking "I agree" you consent to the use of cookies for non-essential functions and the related processing of personal data. You can adjust your cookie and associated data processing preferences at any time via our "Cookie Settings". Please view our Cookie Policy to learn more about the use of cookies on our website.
This website uses cookies to improve your experience while you navigate through the website. Out of these cookies, the cookies that are categorised as ”Necessary” are stored on your browser as they are as essential for the working of basic functionalities of the website. For our other types of cookies “Advertising & Targeting”, “Analytics” and “Performance”, these help us analyse and understand how you use this website. These cookies will be stored in your browser only with your consent. You also have the option to opt-out of these different types of cookies. But opting out of some of these cookies may have an effect on your browsing experience. You can adjust the available sliders to ‘Enabled’ or ‘Disabled’, then click ‘Save and Accept’. View our Cookie Policy page.
Necessary cookies are absolutely essential for the website to function properly. This category only includes cookies that ensures basic functionalities and security features of the website. These cookies do not store any personal information.
Cookie
Description
cookielawinfo-checkbox-advertising-targeting
The cookie is set by GDPR cookie consent to record the user consent for the cookies in the category "Advertising & Targeting".
cookielawinfo-checkbox-analytics
This cookie is set by GDPR Cookie Consent WordPress Plugin. The cookie is used to remember the user consent for the cookies under the category "Analytics".
cookielawinfo-checkbox-necessary
This cookie is set by GDPR Cookie Consent plugin. The cookie is used to store the user consent for the cookies in the category "Necessary".
cookielawinfo-checkbox-performance
This cookie is set by GDPR Cookie Consent WordPress Plugin. The cookie is used to remember the user consent for the cookies under the category "Performance".
PHPSESSID
This cookie is native to PHP applications. The cookie is used to store and identify a users' unique session ID for the purpose of managing user session on the website. The cookie is a session cookies and is deleted when all the browser windows are closed.
viewed_cookie_policy
The cookie is set by the GDPR Cookie Consent plugin and is used to store whether or not user has consented to the use of cookies. It does not store any personal data.
zmember_logged
This session cookie is served by our membership/subscription system and controls whether you are able to see content which is only available to logged in users.
Performance cookies are includes cookies that deliver enhanced functionalities of the website, such as caching. These cookies do not store any personal information.
Cookie
Description
cf_ob_info
This cookie is set by Cloudflare content delivery network and, in conjunction with the cookie 'cf_use_ob', is used to determine whether it should continue serving “Always Online” until the cookie expires.
cf_use_ob
This cookie is set by Cloudflare content delivery network and is used to determine whether it should continue serving “Always Online” until the cookie expires.
free_subscription_only
This session cookie is served by our membership/subscription system and controls which types of content you are able to access.
ls_smartpush
This cookie is set by Litespeed Server and allows the server to store settings to help improve performance of the site.
one_signal_sdk_db
This cookie is set by OneSignal push notifications and is used for storing user preferences in connection with their notification permission status.
YSC
This cookie is set by Youtube and is used to track the views of embedded videos.
Analytics cookies collect information about your use of the content, and in combination with previously collected information, are used to measure, understand, and report on your usage of this website.
Cookie
Description
bcookie
This cookie is set by LinkedIn. The purpose of the cookie is to enable LinkedIn functionalities on the page.
GPS
This cookie is set by YouTube and registers a unique ID for tracking users based on their geographical location
lang
This cookie is set by LinkedIn and is used to store the language preferences of a user to serve up content in that stored language the next time user visit the website.
lidc
This cookie is set by LinkedIn and used for routing.
lissc
This cookie is set by LinkedIn share Buttons and ad tags.
vuid
We embed videos from our official Vimeo channel. When you press play, Vimeo will drop third party cookies to enable the video to play and to see how long a viewer has watched the video. This cookie does not track individuals.
wow.anonymousId
This cookie is set by Spotler and tracks an anonymous visitor ID.
wow.schedule
This cookie is set by Spotler and enables it to track the Load Balance Session Queue.
wow.session
This cookie is set by Spotler to track the Internet Information Services (IIS) session state.
wow.utmvalues
This cookie is set by Spotler and stores the UTM values for the session. UTM values are specific text strings that are appended to URLs that allow Communigator to track the URLs and the UTM values when they get clicked on.
_ga
This cookie is set by Google Analytics and is used to calculate visitor, session, campaign data and keep track of site usage for the site's analytics report. It stores information anonymously and assign a randomly generated number to identify unique visitors.
_gat
This cookies is set by Google Universal Analytics to throttle the request rate to limit the collection of data on high traffic sites.
_gid
This cookie is set by Google Analytics and is used to store information of how visitors use a website and helps in creating an analytics report of how the website is doing. The data collected including the number visitors, the source where they have come from, and the pages visited in an anonymous form.
Advertising and targeting cookies help us provide our visitors with relevant ads and marketing campaigns.
Cookie
Description
advanced_ads_browser_width
This cookie is set by Advanced Ads and measures the browser width.
advanced_ads_page_impressions
This cookie is set by Advanced Ads and measures the number of previous page impressions.
advanced_ads_pro_server_info
This cookie is set by Advanced Ads and sets geo-location, user role and user capabilities. It is used by cache busting in Advanced Ads Pro when the appropriate visitor conditions are used.
advanced_ads_pro_visitor_referrer
This cookie is set by Advanced Ads and sets the referrer URL.
bscookie
This cookie is a browser ID cookie set by LinkedIn share Buttons and ad tags.
IDE
This cookie is set by Google DoubleClick and stores information about how the user uses the website and any other advertisement before visiting the website. This is used to present users with ads that are relevant to them according to the user profile.
li_sugr
This cookie is set by LinkedIn and is used for tracking.
UserMatchHistory
This cookie is set by Linkedin and is used to track visitors on multiple websites, in order to present relevant advertisement based on the visitor's preferences.
VISITOR_INFO1_LIVE
This cookie is set by YouTube. Used to track the information of the embedded YouTube videos on a website.